Several years ago John Waterbury and coworkers [Science (1985) 230:74- 76] reported on the ability of certain marine Synechococcus to swim without apparent benefit of flagella or any other discernible aid. Bianca Brahamsha now tells us she is beginning to get a handle on the question of how they move. Biochemical studies showed that the loss of motility following treatment of cells with proteinase K correlates with the loss of an abundant 120-Kd outer membrane protein. The gene encoding the protein was cloned by a reverse genetics approach, but the sequence gave little indication as to function. After establishing a means of introducing foreign DNA into Synechococcus WH8102, Bianca knocked out the 120-kd outer membrane polypeptide,... and the cells don't swim. They still rotate about their longitudinal axis, but they don't go anywhere, as though they're missing a rudder or something. Of course, this could be an indirect effect -- if they can't make a proper outer membrane, motility components may not be able to insert correctly. Nonetheless, she is excited because for the first time a system is in hand to dissect swimming motility in cyanobacteria.
CONTACT: Bianca Brahamsha, Scripps Institution of Oceanography, Univ. of California-San Diego, La Jolla CA 92093, U.S.A. TEL: 619-534-7505, FAX: 619-534-7313, E-MAIL: BBrahamsha@Ucsd.Edu
============
Alternative respiration (as defined by cyanide-insensitive oxygen consumption) has long been known in cyanobacteria. Guenter Peschek sent in some results from his group that call into question exactly what we mean by "alternative". Using reverse phase HPLC, his group was able to identify heme B (associated with cytochrome b) and lesser amounts of hemes A (associated with aa3-type cytochrome c oxidase) and O from chlorophyll-free cytoplasmic membranes of both unicellular and filamentous cyanobacteria. Heme O was observed only in cultures grown semi-anaerobically. Surprisingly, monospecific antibodies raised against aa3-type cytochrome oxidase (from Paracoccus denitrificans) and against bo3-type quinol oxidase (from E. coli) both recognized the same band on denaturing gels of cytoplasmic membrane protein from cultures, whether or not grown anaerobically. The band comigrated with subunit-I protein of cyanobacterial cytochrome c oxidase. Perhaps heme A and heme O both combine with the same apoprotein, depending on the oxygen-dependent availability of the two hemes. If so, then "aa3-type" and "o3-type" oxidases may represent not "alternative oxidases" in the usual sense of the word but rather alternative hemes. This work has been recently published [Auer et al (1995) Biochem Mol Biol Internatl 37:1173-1185].
CONTACT: Guenter Peschek, Institute of Physical Chemistry, University of Vienna, Waehringerstrasse 42, A-1090 Wien, AUSTRIA, TEL: 43-1-343616, FAX: 43-1-3104597
============
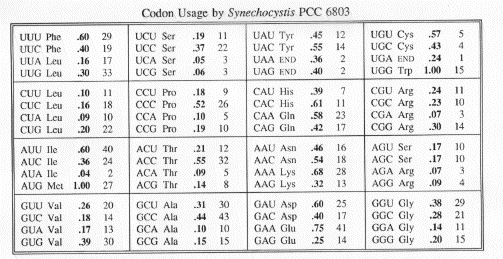
Each codon is followed by a number representing the fraction of instances the amino acid is encoded by the triplet. The second number represents the frequency with which the codon appears on average per 1000 amino acids. The table is based on 10781 codons from sequenced genes of Synechocystis PCC 6803.
CONTACT: Nigel Silman, Biological Sciences, University of Warwick, Coventry, CV4 7AL, U.K., FAX: 203-523701, E-MAIL: Lsrew@Csu.Warwick.Ac.Uk
============
Enumeration of low density cyanobacterial populations is a chancy business, and choices range from the unreliable (plating), to the tedious (microscopy) and horrifically expensive (flow cytometry). Janet Jansson's group recently provided another choice, monitoring luminescence from cyanobacteria tagged with luc, encoding firefly luciferase [Moeller et al (1995) FEMS Microbiol Lett 129:43-50]. Using tagged Synechocystis PCC 6803, they were able to quantitate as few as 4x103 cells per g sediment from a microcosm of Baltic Sea water. Tagged cyanobacteria can also be detected by PCR, taking advantage of the fact that luc is unknown in natural microbial populations, but direct PCR has little quantitative value. Janet now reports that her group has developed a competitive PCR technique that permits quantitation of luc-tagged cyanobacteria in sediment.
The method, soon to be published [Jansson and Lesser (1995) In: Molecular Microbial Ecology Manual, Kluwer Academic Publishers, Dordrecht, Ch. 2.7.4], relies on an internal competitive standard that differs from luc by an additional 35 bp insertion. Otherwise, the target and internal standard are similar and amplified by the same primers during PCR. Using a known concentration of internal standard DNA it is possible to quantitate the original target DNA concentration by the ratio of the amplified products on a gel. The method isn't quite as simple as it sounds, since it is necessary to match the sample to a standard at a comparable concentration, but taking such precautions permitted a very accurate quantitation of luc-tagged Synechocystis in sediment [Moeller and Jansson, manuscript in preparation].
CONTACT: Janet Jansson, Department of Biochemistry, Stockholm University, S-10691 Stockholm, SWEDEN, TEL: 46-8-16-2469, FAX: 46-8-15-3679, E-MAIL: Janet@Biokemi.Su.Se
============
Many of our nonscientific friends know of us vaguely as experts on some sort of algae, and so we are occasionally called upon to identify biological encrustations that we may encounter in daily life. Often we can only respond, lamely, "Looks like green goo to me." Mariona Hernandez-Marine has come to our aid, providing us with a guide to cyanobacteria and algae we might find on buildings and monuments [Ortega-Calvo et al (1995) Sci Total Environ 167:329- 341]. The most commonly encountered cyanobacteria in such places are filamentous species of the genera Phormidium and Microcoleus. We should also point out to our friends that what may appear as very similar blotches of green on the bricks and mortar between them are likely to be quite different communities, owing to a surprising degree of microclimatic variability.
Care must be taken to distinguish such growth from the black sulfated crusts that accumulate on limestone buildings as a result of sulfur dioxide pollution of the air. Even here, however, we must tell our friends that cyanobacteria, particularly of the genus Gloeothece, can thrive on the crusts, despite the presence of toxic compounds, and may provide nutrients for growth of other bacteria.
Some of us may also find ourselves on occasion groping for conversation within dark caves. Here too, Mariona has saved us from a potentially embarrassing assertion, that the grey mat on cave walls could not possibly be due to cyanobacteria. In fact, she tells us, cyanobacteria are amongst the cave's most important epilithic vegetation, despite the very low levels of light. Her description of a calcified cyanophyte of the genus Herpysonema (Mastigocladaceae) has recently been published [Algol Studies (1994) 75:123- 136].
Needless to say, this report only scratches the surface. For further information...
CONTACT: Mariona Hernandez-Marine, Laboratory of Botany, University of Barcelona, E-08028 Barcelona, SPAIN, TEL: 34 3 4024490, FAX: 34 3 4021886, E-MAIL: Hernande@Far.Ub.Es
============
John Smith has alerted us to the fact (pointed out to him by Terry Thiel) that the cyanobacterium he has been calling Nostoc 6720 is more likely to be a species of Anabaena. He confirmed this by comparing the PCR products amplified from genomic DNA, using HIP sequences [Robinson et al (1995) Nucl Acids Res 23:729-735] as primers (courtesy of Nigel Robinson), with those amplified from DNA from Nostoc MAC and Anabaena PCC 7120. Comparison of the nifH sequence from "Nostoc 6720" with the sequence derived by Martin Mulligan from Anabaena variabilis ATCC 29413 suggests that the former strain is closely related to the latter. In fact, stock records identify the cyanobacterium as Anabaena PCC 7937 (nominally identical to ATCC 29413), and the confusion occurred in 1985 when both cyanobacteria were obtained from the Pasteur collection for work on synchronous akinete germination.
CONTACT: John Smith, Dept. of Biological Sciences, University of Lancaster, Bailrigg, Lancaster LA1 3JC, U.K. TEL: 0524-65201, FAX: 0524-843854, E-MAIL: R.Smith@Lancaster.Ac.Uk
============
Many have yearned for the day when the power of bacterial genetics could be applied to the task of elucidating the biosynthetic pathways leading to toxic peptides made by cyanobacteria. The goal has proven to be elusive, however, and until that day arrives, we will have to rely on clever tricks and deductions. Tom Boerner's group at Humboldt University has used both to clone several peptide synthetase genes from Microcystis aeruginosa and to identify a molecular marker for toxic strains.
They exploited the fact that peptide synthetases studied in Gram- positive bacteria and fungi share two highly conserved adenylate-forming domains. Using primers derived from these domains, the Berlin group obtained four different PCR products, two each from DNA of the toxic M. aeruginosa strains HUB 524 and PCC 7820. All four showed striking sequence similarity to peptide synthetase genes. One of the products hybridized to DNA from three tested toxic strains but not to three tested nontoxic strains. That product was used to identify corresponding sequences from genomic libraries, and the 2982 bp sequence of the region (deposited in the EMBL data base with accession number Z28338) showed extended amino acid sequence similarity to peptide synthetases, particularly to the proline-activating synthetase unit of gramicidin S synthetase from Bacillus brevis.
Their results, soon to appear in FEMS Microbiological Letters, indicate that toxic Microcystis may utilize nonribosomal peptide synthesis of the type used to synthesize other peptide toxins. Furthermore, toxic strains may differ from nontoxic strains in part by the presence of a gene or genes required for the synthesis.
CONTACT: Tom Boerner, Institut fuer Biologie, Humboldt-Universitaet, Invalidenstr. 43, D-10115 Berlin, GERMANY, TEL: 49-30-28 97 26 33, FAX: 49-30-28 97 27 99, E-MAIL: Thomas=Boerner@Rz.Hu-Berlin.De
============
Cholera, once confined largely to the Indian subcontinent, has in the last 200 years become a world disease. The disease has swept through much of humanity in a succession of waves, the last major pandemic initiating in 1961. In between incidents of mass infection, Vibrio cholerae, the causative agent, must reside in some still unknown environmental reservoir. It may be pertinent that the peak incidence of cholera in Bangladesh coincides with the appearance of cyanobacterial blooms. Some have postulated that algae and cyanobacteria may in fact constitute the unknown reservoir [Epstein (1993) BioSystems 31:209], pointing to the ability of V. cholerae to survive long periods in the slime produced by these organisms.
Igor Brown has proposed a different connection between cyanobacteria and V. cholerae. He points out that growth of members of the Vibrionacea is stimulated by induction of sodium cycle energetics [Bakeeva et al (1986) Biochim Biophys Acta 850:466]. Brown's own work has suggested that the growth of cyanobacteria in brackish water is autocatalytic: sodium plus alkalinity stimulate the sodium cycle in cyanobacteria, and the resulting growth increases the alkalinity [Brown et al (1990) Biol Membr 4:2039; unpublished results]. The alkalinization and the accompanying increase in dissolved organic compounds resulting from the bloom may induce the sodium cycle in the Vibrionacea, including V. cholerae. Dissemination of the disease would then occur as water from the bloom is used or dispersed.
Whether or not Brown's suggestion is correct, it is comforting to know that cyanobacteria may participate in matters of utmost importance to humans and their funding agencies, quite apart from the prosaic tasks of maintaining the atmosphere and the food chain.
Igor would love to cooperate with anyone in a position to test his idea.
CONTACT: Igor Brown, Cyanobacter Biol. Res. Lab, Odessa State University, Petr Velikiy St. 2, Odessa 270100, UKRAINE, TEL: 007-0482-68-77-93, FAX: 007-0482-23-82-88, E-MAIL: IBrown@Microalgae.Odessa.Ua
============
This past summer Bart Nelissen (U. Antwerpen) defended his doctoral thesis entitled Phylogenetic study of the cyanobacteria on the basis of 16S rRNA gene sequence analysis. Nearly complete 16S rRNA sequences of eleven cyanobacteria belonging to different morphological groups were determined from cloned PCR-amplified products to gain a better understanding of cyanobacterial phylogeny. A cyanobacterium-specific oligonucleotide probe was developed to distinguish cyanobacterial 16S rRNA sequences from amplified products originating from contaminating bacteria. Phylogenetic trees were constructed using the cyanobacterial sequences aligned with other previously determined sequences.
The thesis addressed the phylogenetic relationships between filamentous helical cyanobacteria (Spirulina and Arthrospira) and between cyanobacteria and plastids, and the homogeneity of the genera Pseudanabaena and Leptolyngbya. Most of the results have been published [Nelissen et al (1994) Syst Appl Microbiol 17:206-210; Nelissen et al (1995) Mol Biol Evol 12:1166-1173; Nelissen et al (1996) J Mol Evol in press].
CONTACT: Bart Nelissen, Dept. Biochemie, Universiteit Antwerpen (U.I.A.), Universiteitsplein 1, B-2610 Wilrijk (Antwerpen), BELGIUM, TEL: 32-3-820-23-05, FAX: 32-3-820-22-48, E-MAIL: Nelissen@Uia.Ua.Ac.Be
============
Back to Cyanonews 12-1 Index
Back to Cyanosite
{kind=link}